Birkedal R, Laasmaa M, Vendelin M
Front Physiol 2014;5:376
PMID: 25324784
Full text: http://dx.doi.org/10.3389/fphys.2014.00376
Abstract
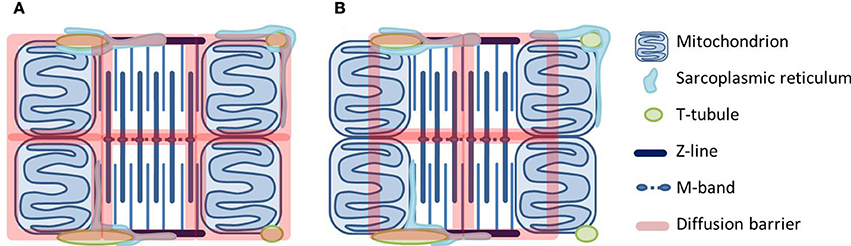
The heart relies on accurate regulation of mitochondrial energy supply to match energy demand. The main regulators are Ca(2+) and feedback of ADP and Pi. Regulation via feedback has intrigued for decades. First, the heart exhibits a remarkable metabolic stability. Second, diffusion of ADP and other molecules is restricted specifically in heart and red muscle, where a fast feedback is needed the most. To explain the regulation by feedback, compartmentalization must be taken into account. Experiments and theoretical approaches suggest that cardiomyocyte energetic compartmentalization is elaborate with barriers obstructing diffusion in the cytosol and at the level of the mitochondrial outer membrane (MOM). A recent study suggests the barriers are organized in a lattice with dimensions in agreement with those of intracellular structures. Here, we discuss the possible location of these barriers. The more plausible scenario includes a barrier at the level of MOM. Much research has focused on how the permeability of MOM itself is regulated, and the importance of the creatine kinase system to facilitate energetic communication. We hypothesize that at least part of the diffusion restriction at the MOM level is not by MOM itself, but due to the close physical association between the sarcoplasmic reticulum (SR) and mitochondria. This will explain why animals with a disabled creatine kinase system exhibit rather mild phenotype modifications. Mitochondria are hubs of energetics, but also ROS production and signaling. The close association between SR and mitochondria may form a diffusion barrier to ADP added outside a permeabilized cardiomyocyte. But in vivo, it is the structural basis for the mitochondrial-SR coupling that is crucial for the regulation of mitochondrial Ca(2+)-transients to regulate energetics, and for avoiding Ca(2+)-overload and irreversible opening of the mitochondrial permeability transition pore.